Podospora anserina Genome Project
- What is Podospora anserina ?
- Where and When can I find Podospora anserina ?
- How does Podospora anserina look like ?
- What is the role of Podospora anserina in nature ?
- How does Podospora anserina reproduce ?
- DoesPodospora anserina age ?
What is Podospora anserina ?
Podospora anserina (Ces.) Rehm is a filamentous fungus that belongs to the class of ascomycetes. It is also known as Pleurage anserina (Ces.) Kuntze. Its complete identification card is as follows: Eukaryota (Cells with a nucleus); Eumycetes (True fungi); Ascomyceta (Sexual spore enclosed within an ascus); Sordariales (Presence of perithecium without a bright color); Lasiosphaeriaceae; Podospora anserina.
Here is a picture that summarises the evolution of eukaryotes as we know it today:
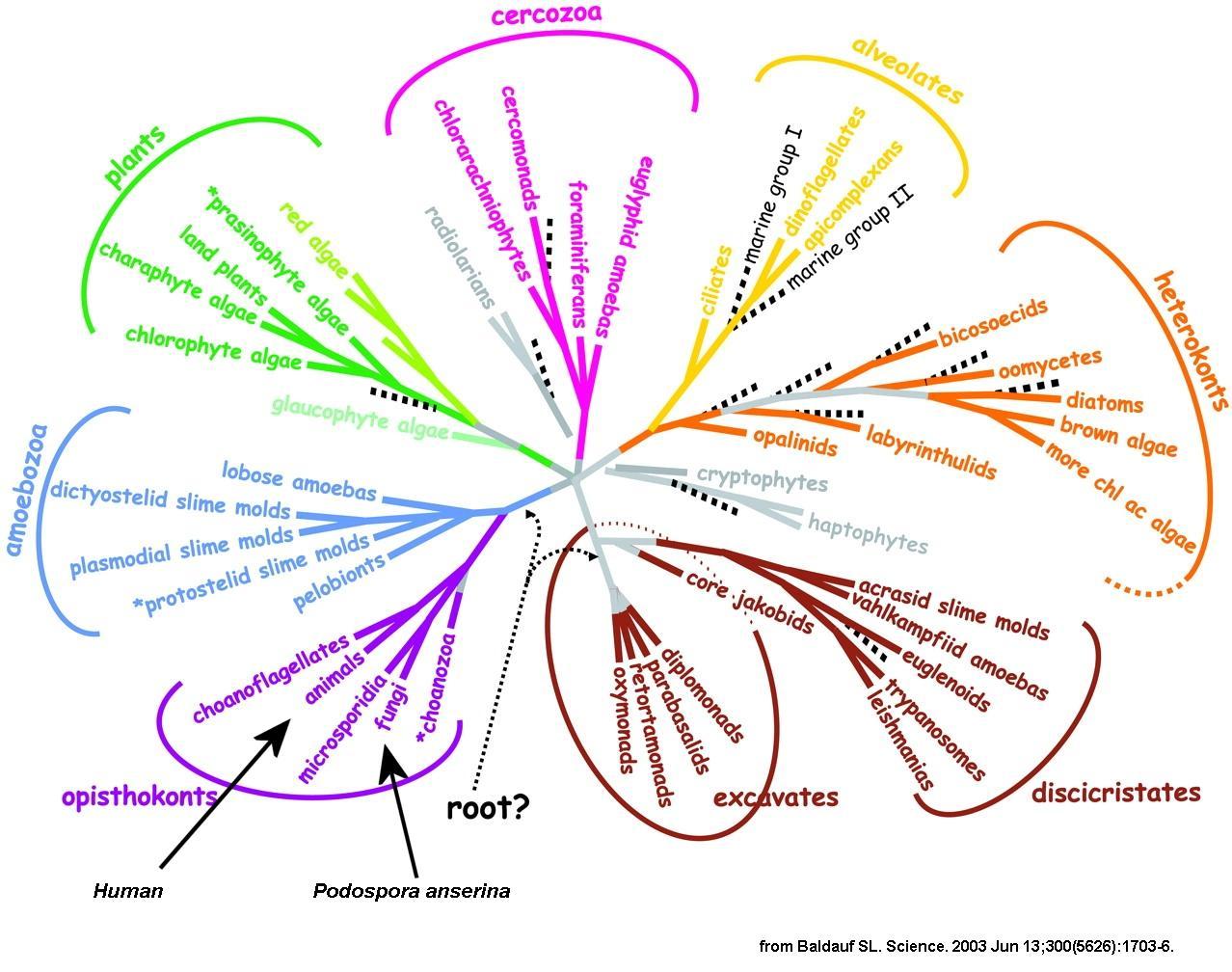 |
Note on this picture that fungi and animals (and thus human) are not as evolutionary distant as is commonly thought. Especially, you can see that plants have diverged before. Among the fungi, it is possible to distinguish several classes. The two principal ones are the Basidiomycetes that includes most of the edible fungi and the Ascomycetes that comprises many different species, including the baker or wine yeast. Truffles and morels are ascomycetous fungi not very distant from Podospora. Neurospora and Sordaria also belong to the Sordariales and are very close relatives. They diverged from Podospora about 75 millions years ago. These three fungi possess a similar life cycle.
If you want to know more on global phylogeny, go to:
If you want to know more on fungi, go to:
If you want to know more about Podospora anserina, check:
Where and When can I find Podospora anserina ?
Podospora anserina lives on and in herbivorous animal dung. It is frequently encountered during summer. The exact geographic distribution of this fungus is unknown. We known that it is commonly found in Western Europe and seems to be present all over the world. If you wish to recover strains from nature in order to grow them in the lab, you need to fetch dung from the fields (horse dung seem rather efficient!) and let the various fungi grow. Indeed, dung from herbivores contains many fungal species that sequentially grow (starting with Mucor-like species, then Sordaria-like species, etc.). P. anserina is one of the last species to grow. It will produce fructifications (=perithecia) that will yield a progeny (=ascospores). Perithecia can be easily spotted on the dung and ascospores can be recovered on an agar plate as described here. To avoid bacterial contamination it is preferable to fetch ascospores directly in the perithecium. Once ascospores are recovered, they can be made to germinate as described here and cultures can be started.
How does Podospora anserina look like?
Like all the filamentous fungi, P. anserina is constituted by hyphae that form a network: the mycelium. These hyphae are made of aligned cells or articles. Only the article located at the end of this line (also called the apical cell) divides. It contains several nuclei, like most cells in the mycelium. Some of the cells are branched allowing for the hyphae to multiply. It is possible to cut the mycelium in small pieces and to regenerate new mycelium from each of these pieces when placed on an appropriate growth medium. Here is a picture of P. anserina hyphae taken under the microscope.
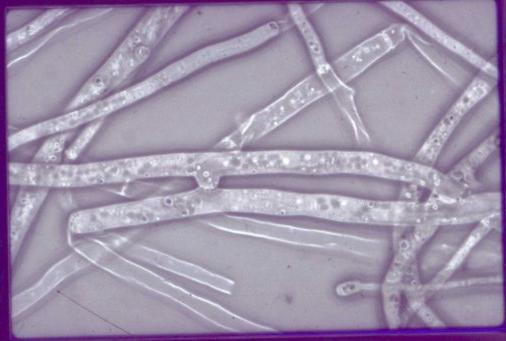 |
At the macroscopic level, the mycelium grows as a disk. However, it can display many different forms, depending upon the growth medium. Some media permit the differenciation of numerous aerial hyphae and the mycelium looks like velvet. Others prevent growth of such hyphae and the mycelium is smooth. Colour also depends upon the growth medium. Most of the time it is greenish but it may also be brownish or the pigments may also be absent.
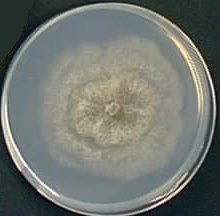 |  | 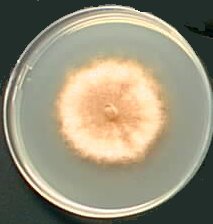 | ||
| M2 Medium | MR Medium | G Medium |
What is the role of Podospora anserina in nature?
Many fungal species have evolved through natural selection life strategies that permit them to use dead plant material as carbon and energy sources. These fungi produce and export outside the cells many enzymes that allow them to degrade plant polymers (like cellulose or lignin). Released nutrients are then transported into the fungal cell by very efficient transport systems. Such nutrition strategy is called saprophytism. P. anserina is such a saprophytic fungi that has specialized to retrieve its nutrients from materials that have not been completely digested by herbivorous vertebrates. It's main function is thus to help to recycle the carbon that is incorporated into highly resistant plant material. This is an important function very often overlooked.
How does Podospora anserina reproduce ?
Podospora anserina is a heterothallic filamentous ascomycete, whose haplobiontic sexual cycle displays interesting steps. Under appropriate conditions (i.e. after exhaustion of the growth medium), mycelia of each mating-type (mat+ and mat-) are able to differentiate male (the microconidia) and female (the ascogonia) "organs". The fertilization event occurs only between a single microconidia and an ascogonium of opposite mating-types. The ascogonium produces the trichogyne, a cytoplasmic extension that "catches" a microconidia. The fertilizing male nucleus enters the ascogonium. An interesting point is that this fertilization event is not immediately followed by nuclear fusion (also called caryogamy). The two nuclei first divide in a common cytoplasm to form a syncitium. The resulting nuclei migrate by pairs of opposite mating type in specialized cells: the ascogenous hyphae. These hyphae give rise, after one or two divisions, to the crozier cells, where the two nuclei divide synchronously. A special septum is formed to yield three cells. Caryogamy usually takes place in the upper binucleate cell. It is immediately followed by meiosis, a postmeiotic mitosis and formation of four binucleate ascospores or of five ascospores - three binucleate and two unicleate ascospores (see genetic section to understand the implication of this ascospore formation process). All these events take place in a fruiting-body structure (the perithecium). The process takes in optimal condition about 4 days at the end of which the ascospores are ejected out of the perithecium by a mechanism that involves turgor pressure.
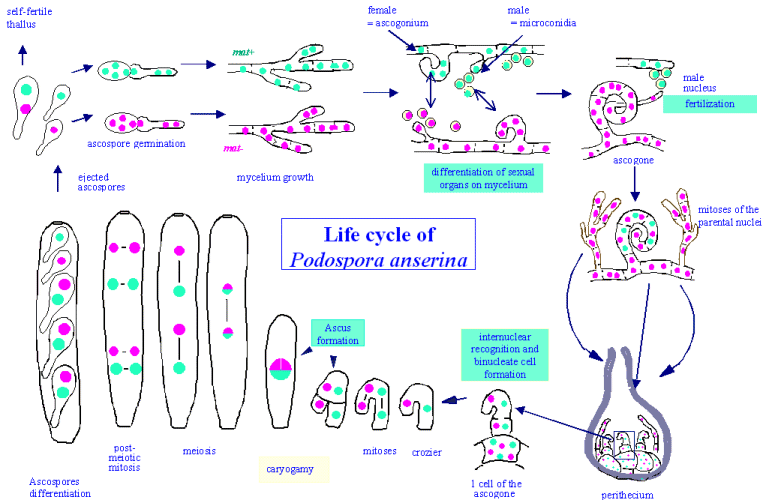 |
Sexual reproduction is the principal mean of dispersion of Podospora as it lacks any special mean of asexual reproduction, especially the fertilizing microconidia do not germinate. However, fragmentation of the mycelium will regenerate new thalli. This is a routinely procedure used in the labs. Note that the transition between a syncytial and a cellular state is a key step during development. In particular, during the evolution of eukaryotes, the appearance of a pluricellular organization remains unsolved. At least two scenarios could explain this transition. In the first one, the pluricellular state is assumed to come from the aggregation of individual cells (as observed in the cycle of the slime mould, Dictyostelium discoideum or the green algae Volvox carteri). The second one assumes that the pluricellular organization arises from a syncitial state. It is illustrated by the early stages of insect development (particularly well described in Drosophila) but also by the transition during the Podospora sexual cycle that leads to the binucleate cells. Thus, the genes, which control this syncitial/cellular switch, are of considerable interest for both developmental and evolutionary points of view. They are intensively searched in Podospora.
Does Podospora anserina age ?
Unlike to many fungi, vegetative growth of P. anserina is not unlimited. It stops definitively at one point and is accompanied by the death of the apical cells. This phenomenon has been called Senescence. Here is a picture of a culture that presents Senescence (on the right part of the culture).
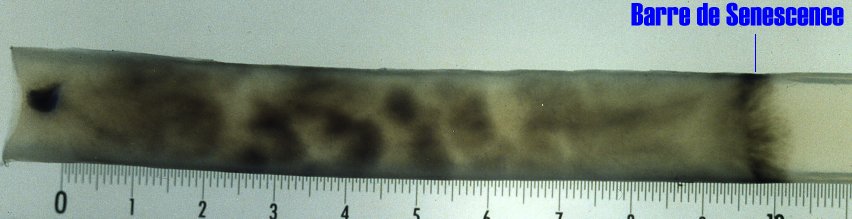 |
On this Podospora culture, ascospore germination occurred on the left. The mycelium then developed toward the right until growth ceased. An intense pigmentation or Barre de Sénescence accompanies the growth stoppage. In this Barre de Sénescence, cells are dead. Cells located closer from the germination point are still alive. This culture grew for about three weeks before dying. The following animation depicts the process in a dynamical manner.
 |
At the cellular level, Senescence is accompanied by a drastic reorganisation of the internal membrane network (mitochondria, endoplasmic reticulum, nuclear membrane). At the molecular level, it is correlated with mitochondrial DNA modifications. It is caused by the accumulation of a cytoplasmic and infectious element, whose nature is still unknown. Longevity can be measured in centimetres of growth! This greatly facilitates measures. Longevity is under the control of numerous nuclear genes (at least 600), the mitochondrial genes, some less defined genetic elements and environmental conditions. This very easy to study aging process is under intense scrutiny in several laboratories in order to identify genes involved in life span definition.